
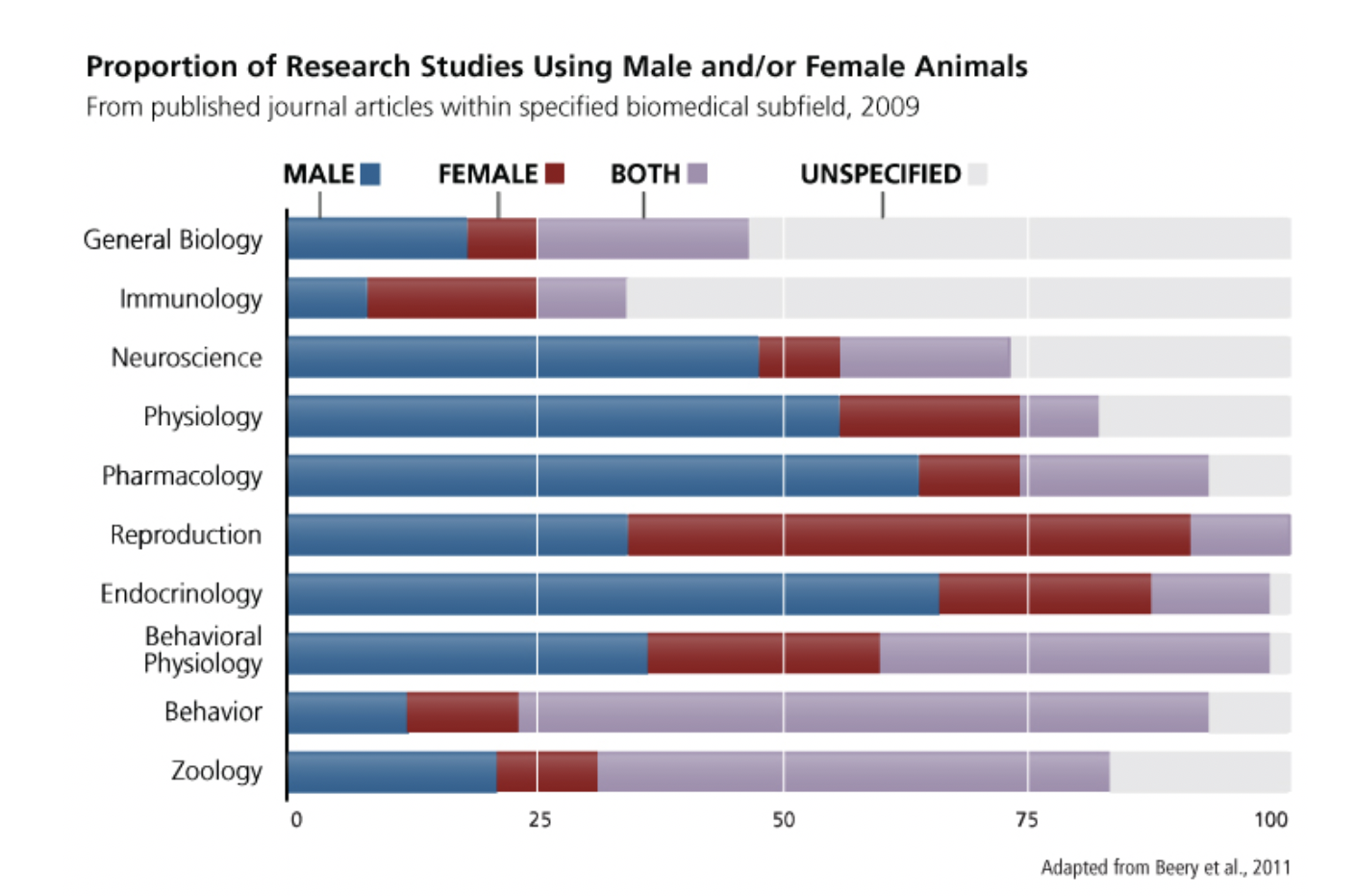
When can experiments be done in only one sex?
- 1. When studying a sex-specific phenomenon, such as ovarian cancer or prostate cancer (McCarthy & Becker, 2002).
- 2. To address inadequate published data for one sex in a particular area. 3. Where there is statistically robust evidence that sex does not influence a trait or outcome.
Sex and Lab Environment in Animal Studies
-
How can we best design animal studies to take into account sex (biological characteristics) and environmental factors and processes? The figure below shows the complex interdependency of sex and environment, including researcher sex, throughout the rodent life cycle.
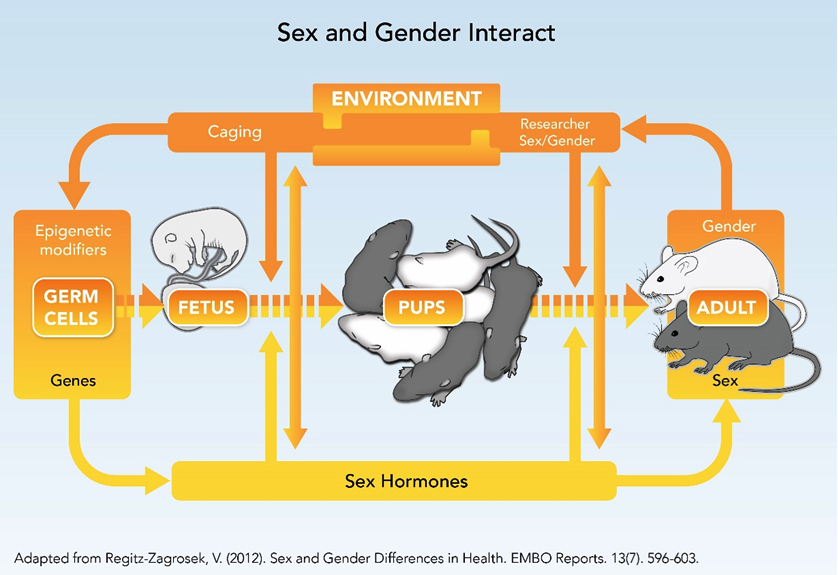
Animal research includes the interaction between sex (biological characteristics, such as genes, hormones, age, reproductive phase, strain, etc.) and extrinsic lab conditions (e.g. caging practices, sex and behaviors of researchers, room temperature, diet, etc.).
Research Design
A. Investigate the Role of Sex
-
• The influence of sex must be investigated before it can be ruled out.
- • Research can be done stepwise. Male and female animals should be strain-matched (or strain-matched and genotype-matched) and age-matched, and should be reared under identical conditions (cages, bedding, diet, etc.). Females should not be breeders unless required for assessment of the phenotype.
- Step 1. Total sample size (based on power calculations). Including females and males in a study often requires doubling the number of experimental subjects, but not always. Efficient experimental designs can incorporate sex while maintaining control over variance (Beery, 2018). For example, factorial designs require sample sizes to be increased by 14–33%, but not doubled (Buch et al., 2019; Miller et al., 2016). Analyzing data by sex and environmental factors helps detect meaningful effects that, in turn, help reduce confounding variables and the cumulative number of experiments required.
- Step 2. Sex-based powering: test hypothesis in both males and females and power each to determine effect. Consider factorial designs to reduce sample size when investigating sex-treatment interactions in animals (Beery, 2018; Lazic, 2018).
- Step 3. Comparison between sexes: power studies to determine the actual “sex effect.” Testing for sex effects has a financial cost. A demonstrated sex difference, however, justifies sex-specific research because harm in one sex is costly to society and individual patients. Overall, it is less expensive to understand sex in the basic science phase than during the more costly clinical trial phase (Klein et al., 2015). Analyzing sex may decrease the number of drugs that fail in development and also helps companies avoid being forced to remove drugs from the market due to adverse events in one sex.
- • Research can be done stepwise. Male and female animals should be strain-matched (or strain-matched and genotype-matched) and age-matched, and should be reared under identical conditions (cages, bedding, diet, etc.). Females should not be breeders unless required for assessment of the phenotype.
- • Researchers should also evaluate overlap between groups (similarities between males and females) and difference within groups (differences among males or among females). Overemphasizing sex differences should be avoided.
- • Finding no sex effect should also be reported. To reduce publication bias, researchers should report when sex differences (main effects or interaction effects) are not detected or when data regarding sex differences are statistically inconclusive (Wizemann, 2012). Reporting null results is crucial for meta-analysis.
- • For phenotypes that do not display sex difference, future experiments should be sex inclusive; that is, they should include equal numbers of randomly selected males and females for each test group studied. Not every experiment needs to be designed to evaluate sex differences. However, for every experiment, the sex of the animal test subjects should be reported to ensure that experiments are reproducible and findings in one sex are not over-generalized to the other sex (Wizemann, 2012).
- • Analyze the Impact of Sex Hormones
- Considering the estrous cycle (Byers et al., 2012). Research has shown that the majority of sex effects are not influenced by cycle (Becker et al., 2005). Researchers should compare the variability in female and male responses. Researchers do not need to monitor the estrus cycle unless females are more variable.
- • In a meta-analysis of nearly 10,000 traits, Prendergast et al. (2014) found that, for most biological measurements, females are no more variable than males (Itoh & Arnold, 2015; Becker et al., 2016). Other factors, including group versus single animal housing, can have a greater impact on variability of a trait than stages of the estrous cycle.
- Menopause Models. Menopause is an emerging area of laboratory animal research. In rodents, reproductive aging occurs and can be used in research to gain insight into menopause in women. Reproductive aging in female rats or mice begins towards the end of the first year, and the timing can vary depending on strain and environmental factors (Finch et al., 1984). When over one year old, female rodents show either low serum levels of gonadal steroids (persistent diestrus) and/or chronically elevated serum estrogen levels (persistent estrus) (Finch et al., 1984). Persistent estrus leads eventually to a diestrus condition, but there can be an intervening period of pseudopregnancy, depending on the species (Finch et al., 1984). Ovariectomy can simulate reproductive aging by rapidly reducing gonadal steroid levels, and this “acute menopause” has a multitude of effects. One area of increasing interest is the effects of menopause on the immune system. In rodents, ovariectomy “reduce[s] lymphocyte chemotaxis, mitogen-induced T cell proliferation responses, and [Interleukin-2] production” (Marriott & Huet-Hudson, 2006).
- Pregnancy or Pseudopregnancy. Fewer than 10% of medications approved by the U.S. Food and Drug Administration since 1980 have enough information to determine risks for birth defects (Adam et al., 2011; Mishra & Mohanty, 2010). New animal research on drug safety should assess effects on the dam and the fetus during pregnancy and lactation (McDonnell-Dowling & Kelly, 2015).
- Pharmacokinetics. The estrous cycle can also affect pharmacokinetics. For example, Kulkarni et al. (2012) found that the oral bioavailability of genistein, a soy isoflavone with antioxidant properties, was inversely correlated with estrogen level (which regulates hepatic disposition of a drug).
- Considering the estrous cycle (Byers et al., 2012). Research has shown that the majority of sex effects are not influenced by cycle (Becker et al., 2005). Researchers should compare the variability in female and male responses. Researchers do not need to monitor the estrus cycle unless females are more variable.
-
B. Investigate How the Environment Interacts with Sex
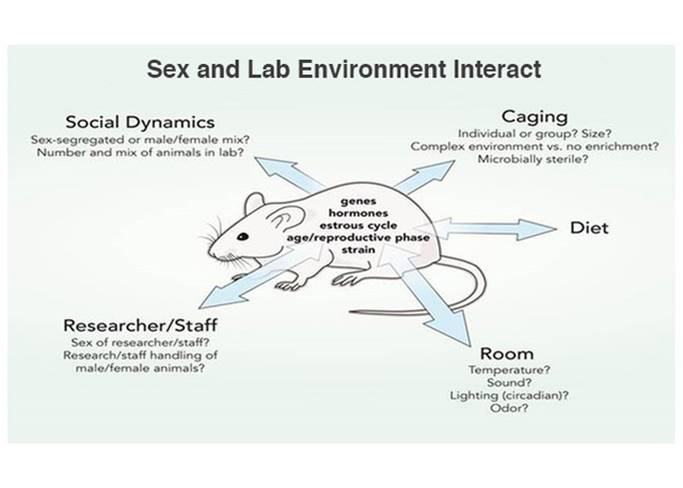
Animal research includes the interaction between sex (biological characteristics, such as genes, hormones, age, reproductive phase, strain, etc.) and the lab environment (e.g. caging practices, attitudes and behaviors of researchers, room temperature, diet, etc.). The double-ended arrows represent interactions between sex and environment. Environmental processes, such as caging practices or differential handling, may impact female and male animals differently. Researchers should not identify an effect as dependent on sex (or a biological trait) when, in fact, it depends on an environmental condition.
Environmental Processes that May Interact with Sex:
- Caging: Individual vs. group?
- • To avoid aggressive behaviors, male rodents are often caged in small groups or alone. Rodents housed alone “expend more energy maintaining body temperature, which can cause differences in parameters such as caloric intake, muscle activity, metabolic rate, fat distribution, or body size, with a plethora of potential downstream effects on bodily and cellular activity” (Ritz et al., 2014). In contrast, females are more often housed together to lower costs. Rodents housed together often sleep clustered and, as a result, expend less energy to keep warm. In this scenario a “sex difference” may be identified where, in fact, differences result from different housing conditions.
- • A same-sized group may create different stressors for females and males. Being caged alone may itself cause stress (Ritz et al., 2014), but single housing reduces trait variability in both males and females (Prendergast et al., 2014).
- • Group caging can also result in self-induced or social hair loss, also called barbering (Kaleuff et al., 2006). Barbering (1) often reflects social hierarchies in same-sex group cages (both females and males); (2) may result from the stress of overcrowding; (3) takes place in breeding groups (females barber males); and (4) occurs among lactating rodents (pups barber mothers). Barbering occurs in some strains more than others.
- • Cage size can limit animal behavior. For example, many cages cannot accommodate the full range of female sexual behavior. In the wild, females may dart, approach and solicit males (Birke, 2011). For these reasons, articles should specify housing conditions.
- Researcher/Staff. Experimenters may be a confounding variable in rodent research where stress is a significant factor. One study found that rats and mice demonstrated a reduced pain response in the presence of a male experimenter compared to a female experimenter. Both male and female rodents showed this response, but females showed it more. The researchers identified this “male observer effect” as a stress response to androstenone and androstadienone, axillary secretions found in higher concentrations in males than females. In addition to stress-induced analgesia, the presence of these compounds resulted in increased plasma corticosterone levels (Sorge et al., 2014).
- Handling. Control rodents should undergo similarly stressful procedures as those of the experimental rodents, such as sham surgeries. Taking vaginal smears to establish the stage of the estrous cycle in female rodents can be stressful; male rodents should be handled in similar ways (Becker et al., 2005). Without these controls differences in stress responses can be mistaken for other sex differences.
- Circadian Cycling. Testosterone varies seasonally and with circadian rhythms. Similarly, hormone concentrations in females can fluctuate over the course of a single day of the estrous cycle. Hypothalamo-pituitary-adrenal (HPA) secretions, which in turn affect gonadal secretions, also vary over the course of the day (Becker et al., 2005). Researchers should specify the photoperiod in the colony and the time of day at which measurements are taken.
- Social Dynamics. Edelmann et al. (2013) found that rat maternal behavior mediates sex differences in play among juveniles. Simulated maternal grooming, in addition to normal maternal care, reduced play in males but not in females. This effect may be mediated by increased serotonin signaling, as maternal licking also increased serotonin receptor mRNA.
- Temperature. Laboratory mice are typically housed at temperatures below their thermoneutral zone. Gaskill et al. (2009) found that when mice were able to move among three cages with different temperatures, mice of both sexes preferred warmer environments for inactive and maintenance behaviors (with no preference for active behaviors). Females preferred the highest temperature; males showed no preference between the medium and highest temperatures. As with day length, ambient temperatures in the research colony should be reported consistently.
- Diet. Diets impact weight, metabolism, hormone levels and immune functions; hence diet formulation should be reported (Bhupathy et al., 2010; Luczak et al., 2011). Glover & Assinder (2006) found that diets rich in phytoestrogens may have sex-specific effects on cardiac health. In males, soy-based diets significantly decreased cardiac function, increased myocellular disarray and led to an increase in β-MyHC, a myosin motor protein associated with heart failure. This latter effect was also observed to a lesser degree in females.
- Bedding. Bedding might also have an impact on hormonal status. Corncob bedding has been reported to reduce aggressive behavior compared to cardboard-based bedding by influencing estrogen receptor alpha expression in the brain (Landeros et al., 2012). Elevated levels of non-estrogenic mitogens have also been previously reported in mice housed on corncob bedding (Markaverich et al., 2002).
Works Cited
Adam, M., Polifka, J., & Friedman, J. (2011). Evolving knowledge of the teratogenicity of medications in human pregnancy. American Journal of Medical Genetics, Part C., 157, 175-182.
Arrowsmith, J. (2011). Trial watch: phase ii failures: 2008–2010. Nature Reviews Drug Discovery, 10, 328-329. Becker, J. B., Prendergast, B. J., & Liang, J. W. (2016). Female rats are not more variable than male rats: a meta-analysis of neuroscience studies. Biology of sex differences, 7(1), 34. Beery, A. K. (2018). Inclusion of females does not increase variability in rodent research studies. Current Opinion in Behavioral Science, 23, 143–149. Beery, A., & Zucker, I. (2011). Sex bias in neuroscience and biomedical research. Neuroscience and Biobehavioral Reviews, 35(3), 565-572. Buch, T., Moos, K., Ferreira, F. M., Fröhlich, H., Gebhard, C., & Tresch, A. (2019). Benefits of a factorial design focusing on inclusion of female and male animals in one experiment. Journal of Molecular Medicine, 97(6), 871-877. Bhupathy, P., Haines, C., & Leinwand, L. (2010). Influence of sex hormones and phytoestrogens on heart disease in men and women. Women's Health 6(1), 77–95. Birke, L. (2011). Telling the rat what to do: laboratory animals, science, and gender. In Fisher, J. A. (Ed.), Gender and the Science of Difference; Cultural Politics of Contemporary Science and Medicine (pp. 91-107). New Brunswick: Rutgers University Press. Byers, S., Wiles, M., Dunn, S., & Taft, A. (2012). Mouse estrous cycle identification tool and images. PLoS ONE, 7(4). doi:10.1371/journal.pone.0035538. DiMasi, J. A., Grabowski, H. G., & Hansen, R. W. (2016). Innovation in the pharmaceutical industry: new estimates of R&D costs. Journal of Health Economics, 47, 20–33. Edelmann, M. N., Demers, C. H., & Auger, A. P. (2013). Maternal touch moderates sex differences in juvenile social play behavior. PLoS ONE, 8(2). doi:10.1371/journal.pone.0057396 Finch, C. E., Felicio, L. S., Mobbs, C. V., & Nelson, J. F. (1984). Ovarian and steroidal influences on neuroendocrine aging processes in female rodents. Endocrine Reviews, 5(4) 467-497. Gaskill, B. N., Rohr, S. A., Pajor, E. A., Lucas, J. R., & Garner, J. P. (2009). Some like it hot: mouse temperature preferences in laboratory housing. Applied Animal Behavioral Science, 116, 279-285. Glover A., & Assinder S. J. (2006). Acute exposure of adult male rats to dietary phytoestrogens reduces fecundity and alters epididymal steroid hormone receptor expression. Journal of Endocrinology, 189(3), 565-573. Herper, M. (2013, August 11). The cost of creating a new drug now $5 billion, pushing big pharma to change. Forbes. Itoh, Y., & Arnold, A. P. (2015). Are females more variable than males in gene expression? Meta-analysis of microarray datasets. Biology of Sex Differences, 6(1), 18. Kaleuff, A., Minasyan, A., Keisala, T., Shah, Z., & Tuohimaa, P. (2006). Hair barbering in mice: implications for neurobehavioral research. Behavioral Processes, 71(1), 8-15. Klein, S. L., Schiebinger, L., Stefanick, M. L., Cahill, L., Danska, J., De Vries, G. J., ... & Zucker, I. (2015). Opinion: sex inclusion in basic research drives discovery. Proceedings of the National Academy of Sciences, 112(17), 5257-5258. Kulkarni K. H., Yang, Z., Niu, T, & Hu, M. (2012). Effects of estrogen and estrus cycle on pharmacokinetics, absorption, and disposition of genistein in female Sprague-Dawley rats. Journal of Agricultural and Food Chemistry, 60, 7949-7956. Landeros, R. V., Morisseau, C., Yoo, H. J., Fu, S. H., Hammock, B. D., & Trainor, B. C. (2012). Corncob bedding alters the effects of estrogens on aggressive behavior and reduces estrogen receptor-α expression in the brain. Endocrinology, 153(2), 949-953. Lazic, S. E. (2018). Four simple ways to increase power without increasing the sample size. Laboratory animals, 52(6), 621-629. Luczak, E. D., Barthel, K. K., Stauffer, B. L., Konhilas, J. P., Cheung, T. H., & Leinwand, L. A. (2011). Remodeling the cardiac transcriptional landscape with diet. Physiological genomics, 43(12), 772-780. Markaverich, B., Mani, S., Alejandro, M. A., Mitchell, A., Markaverich, D., Brown, T., ... & Faith, R. (2002). A novel endocrine-disrupting agent in corn with mitogenic activity in human breast and prostatic cancer cells. Environmental health perspectives, 110(2), 169-177. Marriott, I., & Huet-Hudson, Y. (2006). Sexual dimorphism in innate immune responses to infectious organisms. Immunologic Research, 34(3), 177-192. McCarthy, M., & Becker, J. (2002). Neuroendocrinology of sexual behavior in the female. In Becker, J., Breedlove, S., Crews, D., & McCarthy, M. (Eds.), Behavioral Endocrinology (pp. 124-132). Cambridge: MIT Press. McDonnell-Dowling, K., & Kelly, J. (2015). Sources of variation in the design of preclinical studies assessing the effects of amphetamine-type stimulants in pregnancy and lactation. Behavioural Brain Research, 279, 87-99. Mehta, L. S., Beckie, T. M., DeVon, H. A., Grines, C. L., Krumholz, H. M., Johnson, M. N., ... & Wenger, N. K. (2016). Acute myocardial infarction in women: a scientific statement from the American Heart Association. Circulation, 133(9), 916-947. Miller, L. R., Marks, C., Becker, J. B., Hurn, P. D., Chen, W. J., Woodruff, T., ... & Makris, S. (2016). Considering sex as a biological variable in preclinical research. The FASEB Journal, 31(1), 29-34. Miller, V. M., Kaplan, J. R., Schork, N. J., Ouyang, P., Berga, S. L., Wenger, N. K., ... & Taylor, D. A. (2011). Strategies and methods to study sex differences in cardiovascular structure and function: a guide for basic scientists. Biology of Sex Differences, 2(1). doi: 10.1186/2042-6410-2-14 Mishra, A. & Mohanty, B. (2010). Effect of lactational exposure of olanzapine on body weight of mice: a comparative study on neonates of both the sexes during post-natal development, Journal of Psychopharmacology, 23(7), 1089-1096. Ouyang, P., Wenger, N. K., Taylor, D., Rich-Edwards, J. W., Steiner, M., Shaw, L. J., ... & Merz, N. B. (2016). Strategies and methods to study female-specific cardiovascular health and disease: a guide for clinical scientists. Biology of Sex Differences, 7(1), 19. Prendergast, B. J., Onishi, K. G. & Zucker, I. (2014). Female mice liberated for inclusion in neuroscience and biomedical research. Neuroscience and Biobehavioral Reviews, 40, 1–5. Regensteiner, J. G., Golden, S., Huebschmann, A. G., Barrett-Connor, E., Chang, A. Y., Chyun, D., ... & Reusch, J. E. (2015). Sex differences in the cardiovascular consequences of diabetes mellitus: a scientific statement from the American Heart Association. Circulation, 132(25), 2424-2447. Ritz, S., Antle, D., Côté, J., Deroy, K., Fraleigh, N., Messing, K., Parent, L., St-Pierre, J., Vaillancourt C., & Mergler, D. (2014). First steps for integrating sex and gender considerations into basic experimental biomedical research. The FASEB Journal, 28(1), 4-13. doi: 10.1096/fj.13-233395. Sorge, R., Martin, L., Isbester, K., Sotocinal, S., Rosen, S., Tuttle, A., Wieskopf, J., Acland, E., Dokova, A., Kadoura, B., Leger, P., Mapplebeck, J., McPhail, M., Delaney, A., Wigerblad, G., Schumann, A., Quinn, T., Frasnelli, J., Svensson, C, Sternberg, W., & Mogil, J. (2014). Olfactory exposure to males, including men, causes stress and related analgesia in rodents. Nature Methods, 11, 629–632. Wizemann, T. (Ed.) (2012). Sex-Specific Reporting of Scientific Research: A Workshop Summary. Washington, D.C.: National Academies Press.



